
Y-kromoszóma: Az embernek 46 kromoszómapárja van. 22 pár testi és egy pár nemi kromoszóma. A nőkre az XX, a férfiakra az XY jellemző. Az Y–kromoszómát csak az apa adhatja át fiú utódjának.
Mitokondrium: Olyan sejt szervecske, mely a sejtek energiatermelésében játszik szerepet. DNS-ük csak anyai ágon öröklődik. Egy átlagos sejtben több száz mitokondrium található, mindegyikükben 5-10 DNS molekula.
Haplotípus: Y-kromoszóma (vagy mitokondrális DNS) típus, melyek meglétét vagy hiányát mérni és ezáltal összehasonlítani lehet a különböző népességék egyedeiben.
Polimorfizmus (változatosság): Egy gén változatból, más néven allélból, a populáción belül legalább kettő van jelen.
Filogenetikus: Fejlődéstörténeti, törzsfejlődési, származástani; a törzsfejlődéssel kapcsolatos.
Paleolitikum (őskőkor): Az emberiség történetének legrégebbi és leghosszabb szakasza. Három nagy korszakra osztható, ebből az utolsó az ún. felső-paleolitikum (késői őskőkor) 35 ezertől 10 000-8000 ig tartott, melynek legfontosabb kultúrája az aurignaci kultúra.
Neolitikum (újkőkor): Az emberiség történetének kb. i.e. 5000-3000-ig tartó időszaka. Ekkor alakult ki a maihoz hasonló éghajlat, állat- és növényvilág. Megkezdődött az állatok háziasítása, tenyészése kialakult a kapás földművelés, háttérbe szorul a vadászat. (Falu jellegű telepek, anyajogú társadalom, nádból, fából és agyagból készített házak.)
Aurignac-i régészeti kultúra: A kései paleolitikum kezdetén virágzott. A párhuzamos élű, pengeszerű kőeszközök és a finoman megmunkált csonteszközök a fő jellemzői.
Gravetti régészeti kultúra: Az aurignac-i kultúrával együtt a felső-paleolitikum másik kiemelkedő kultúrája. Szintén kelet felől (talán a mai Ukrajna) területéről áramlott be és párjával együtt a parányi kőpengéből készített fegyverek, eszközök jellemzik.
Kurgán kultúra: (más néven okker vagy gödörsíros kultúra) Marija Gimbutas, W. P. Lehmann és sok más kutató a sztyeppei kurgán-kultúra területét tekinti indoeurópai őshazának, népét a még differenciálatlan indoeurópai alapnyelv beszélőinek. A Volga és a Dnyeper vidékéről elköltöző nomád, patriarchális és erősen hierarchizált társadalmú népesség elsősorban pásztorkodással foglalkozott.
Az elemzés legfőbb és talán legmeglepőbb konklúziója az volt, hogy a 25 európai és közel-keleti régióból származó több mint ezer fős Y-kromoszóma mintázatok több mint 95%-ban 10 haplotípusba voltak besorolhatók, a teljes 22-ből (1. ábra, felső ágrajz). Semino és munkatársai gondolatmenete szerint a haplotípusok gyakoriságának megoszlása felfedi a modern európai népesség férfi komponenseinek alap összetételét, és bizonyító adatokat szolgáltat a kontinens paleolitikum időszakából indított benépesüléséhez, népességtörténetéhez.
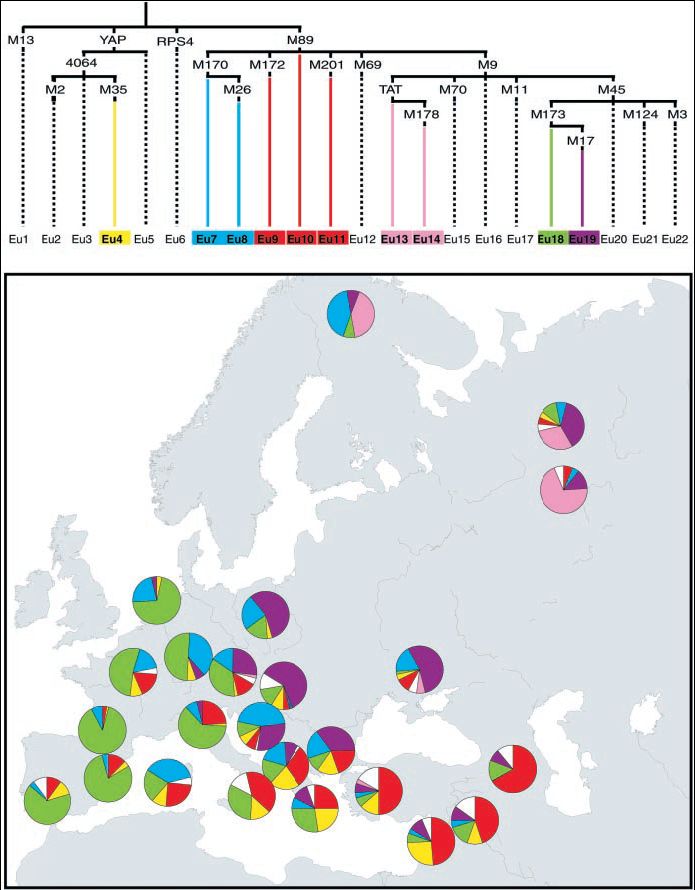
1. ábra (felül) Az Európában talált Y-kromoszóma markerek egyszerűsítet filogenetikus (leszármazási) fája. Az ág elágazási pontjaiban az egyes haplotípusokat definiáló kulcsmutációk, az alsó sávban maguk a haplotípusok (a mutációk azonosító elnevezései) láthatók (a jelentősebbek és egy csoportba sorolandók színekkel kiemelve, megkülönböztetve).
(alul) A jelentősebb haplotípusok földrajzi megoszlása a filogenetikus fa haplotípus ábrázolása szerint. (Az ábra forrása: Semino és mtársai 2000.)
Az eredmények szerint két leszármazás (az M173-as és M17-es mutációkkal jellemezhetőek) van jelen Európában a paleolitikus időkből. A két kromoszómatípus - melynek az Eu18 és az Eu19 nevet adták - körülbelül 50%-ban jellemzi az európai férfiakat, mégpedig akképpen, hogy a két haplotípus egymással ellentétes földrajzi megoszlást mutat a kontinensen belül(1. ábra, alsó kép). Az Eu18 gyakorisága (zöld színnel jelölve) fokozatosan csökken nyugatról kelet felé haladva, leggyakoribb a baszkok között. Ellenben az Eu19 haplotípus (lila színnel jelölve) - mely szintén az M173-as mutációval jelölt leszármazásból ered -, gyakorlatilag hiányzik Nyugat-Európában. Gyakorisága fokozatosan növekszik kelet felé, maximumát Lengyelországban, Magyarországon illetve Ukrajnában éri el, ahol gyakorlatilag az Eu18 hiányzik. Ezen megfigyelések Seminoék szerint felvetik, hogy az M173 egy olyan ősi eurázsiai marker, amit az Európába belépő Homo sapiens sapiens vagy magával hozott, vagy ideérkező csoportjában alakulhatott ki, amelyek szétszóródva elterjesztették ezt a génnyalábot 35-40 ezer évvel ezelőtt. Valószínűsítik, hogy a hordozói az ún. aurignaci régészeti kultúra emberét jelentik.
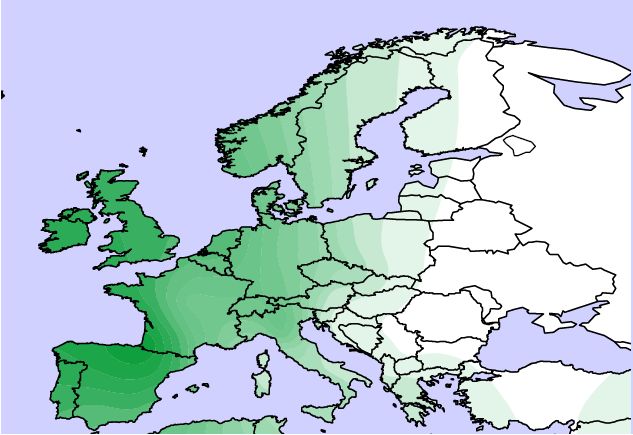
2/b. ábra (felül) Az Eu18-es haplotípus génfrekvencia térképe Európában Semino és mtsai közleménye alapján. (Ábra Gáspár R.)
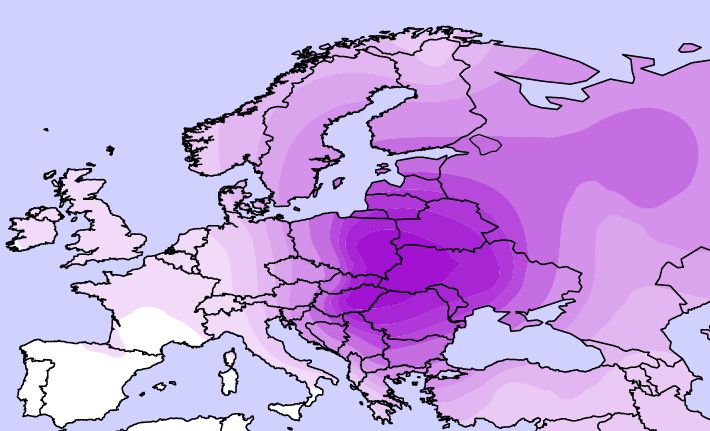
(alul) Az Eu19-es haplotípus génfrekvencia térképe Európában Semino és mtsai közleménye alapján. (Ábra Gáspár R.)
A genetikusok úgy értelmezték az Eu18 és a később ebből újabb egymutáció (M17) által derivált Eu19 haplotípus kontinensen belül tapasztalt megoszlását, hogy ezek mintegy bélyegei a jégkorszak idején az Ibériai-félszigetre és a jelenlegi Ukrajna területére visszahúzódott egymástól hosszú időre elszigetelt népességeknek, illetve jégtakaró visszahúzódását követően expanziójuknak. A jégkorszak alatt (13-20 ezer évvel ezelőtt) a különböző humán csoportok arra kényszerültek, hogy az észak-balkáni menedéket kivéve elhagyják Közép-Európa területét. Hasonlóan az európai növény és állatvilágban található párhuzamokhoz, az eljegesedésnek tulajdonított izolációt ismételt szétszóródás (expanzió) követte a klimatikus menedékekről kiindulva (Ibériai-félsziget, Ukrajna, Észak-Balkán) időszakban. Elterjedését Semino-ék szerint felerősíthette az egykori Kurgán kultúra területéről (Dél-Ukrajna területe napjainkban) kiinduló Yamana régészeti kultúra expanziója Európa illetve kelet felé, magában hordozva az indoeurópai nyelv szétterjedését.
Az Eu19 haplotípussal kapcsolatosan megemlítendő, hogy az nem csak Európában fordul elő. Tekintélyes frekvenciákkal találkozhatunk Észak-Indiában, Pakisztánban és Közép-Ázsiában is, ahol egyébként a marker legnagyobb gyakoriságát éri el. A Yamna kultúra összekapcsolása az indo-európai nyelvvel elterjedésével mint munkahipotézis fogadható el, hiszen erre közvetlen bizonyíték természetesen nem létezik.
Megemlítendő, hogy más kutatók eltérő eredményekhez jutottak a mutáció életkorát tekintve, ami teljesen új megvilágításba helyezheti az Eu 19-es haplotípusról kialakított képet. Három különböző kutató lényegesen későbbire becsülte az M17 életkorát: Zerjal 3800, Kayser 2550 illetve Karafet 7500 évesnek datálta. Ha ezek az adatok tükrözik a valóságot, akkor a magyarokban is nagy gyakorisággal meglévő marker expanziója csakis a Kurgán kultúra vándorlását követő népmozgásokhoz kapcsolható. Rosser alternatív magyarázata alapján inkább a nagyállattartó nomadizmus elterjedéséhez, azaz szkíták, mongolok és a hunok kelet-nyugati vándorlásaihoz köthető.
Az M170-es polimorfizmus egy másik vélelmezett paleolitikumi mutációt mutat, aminek a becsült kora 22 ezer év. Újabb keletű génáramlásra utal, jelenléte Európára korlátozódik túlnyomórészt (Eu7, majd belőle az Eu8). Ezek leggyakoribbak Közép-Kelet Európában, de feltűnik a baszkoknál és a szardíniaiaknál is (1. ábra). Az M170 legközelebbi filogenetikus előde az M89 mutáció, amiből a legfontosabb közel-keleti mutációk is származnak. Ezek alapján feltételezhető, hogy az európai eredetű M170 azoknak a leszármazottaiban található, kiknek ősei a Közel-Keletről indultak el mintegy 20-25 ezer évvel ezelőtt, és elterjesztették a gravetti régészeti műveltség emlékeit.
A gravetti és az aurignaci csoportok több évezredes békés egymás mellett élését csak az eljegesedés korszaka szakította meg, amikor is Nyugat-Európa elszigetelődött Közép-Európától, ahol azonban az epi-gravetti kultúra tovább folytatódott Ausztria, Csehország és az Észak-Balkán régióiban. A klíma kedvezőbbre fordulása után ez a kultúra észak és kelet felé terjedt tovább, mely jelenség párhuzamba állítható az Eu7 haplotípus eloszlásával. Ezen forgatókönyv szerint az Eu8 haplotípus a nyugati paleolitikumi populációkban alakult ki az eljegesedés maximumának időszaka alatt, mint a helyi differenciálódása az M170-es leszármazásnak.
Ezt a felismerést a későbbi vizsgálatok is megerősítették a modern magyar népesség tekintetében. A honfoglalóknál hét mintából kettő esetben tudták kimutatni ezt a bizonyos Tat markert, ami uráli, vagy ha a jelenleg rendelkezésünkre álló archeogenetikai adatokat veszünk figyelembe, Belső-Ázsiába az ázsiai hunok irányába mutathat.
Először Cavalli-Sforza a Stanford Egyetem genetikusa próbálkozott a különböző populációk és nyelvcsaládok között fennálló lehetséges kapcsolatok feltérképezésére. Megállapítása szerint „a genetikai és nyelvi törzsfák közt mégis vannak eltérések, és ezek az eltérések sok forrásból táplálkoznak. Egy nyelvet viszonylag rövid időn belül le lehet cserélni egy másikkal". Tehát a genetikai és nyelvi törzsfák összevetése csak egyes esetekben hozott (hozhat) párhuzamos eredményt, mivel a nyelvek változási sebessége nagyságrendekkel gyorsabb, mint maga a genetikai evolúció, illetve az esetlegesen meglévő kapcsolatot felboríthatja a „nyelvcsere" jelensége. Ezért sem meglepő eredmény, hogy a magyar és a többi nyelvrokon populációk összehasonlítása során a nyelvi (uráli nyelvcsalád tagja) és a genetikai evolúció („uráli" TAT marker hiánya) nem mutat párhuzamot a két vizsgált csoport esetében.
A genetikai kutatások során érdemes tehát a megszülető eredményeket összevetni a különböző tudományágakkal – mint például Seminoék tették jelen közleményben a régészettel –, de azok természetesen nem biztos, hogy azonos irányba fognak mutatni.
Az M35 (Eu4), M172 (Eu9), M89 (Eu10) és M201 (Eu11) haplotípusok gyakorisága Közel-Keleten a legmagasabb, Európa felé haladva fokozatosan csökken frekvenciája (1. ábra). Az Eu4, Eu9 és Eu10 a becsült életkora 15-20 ezer, míg az Eu11 17 ezer évre tehető. Ezek alapján nem állapítható meg, hogy vajon az eljegesedés előtt, vagy után érkeztek a kontinensre, mindazonáltal az Eu4, Eu9, Eu10 és Eu11 hapoltípusok Közel-Kelet felől Európa irányába megfigyelt csökkenő gyakorisága és térbeli elhelyezkedése alapján nem mutatható ki az Eu18 és Eu 19 esetében tapasztalt földrajzi megoszlás. Így - a szerzők szerint - az Eu4, Eu9, Eu10 és az Eu11 a közel-keleti eredetű, földművelést elterjesztő populációk genetikai hozzájárulása lehet Európa férfi génállományához, ami úgy tűnik sokkal hangsúlyosabb a Mediterráneum partvidékén, mint Közép-Európában. Ez leolvasható a 2. ábráról, melyen grafikusan került ábrázolásra az európaiakban kimutatott Eu4, Eu8, Eu10 és Eu11
gyakoriságok, a Közel-Kelet térségétől való földrajzi távolsághoz viszonyítva.
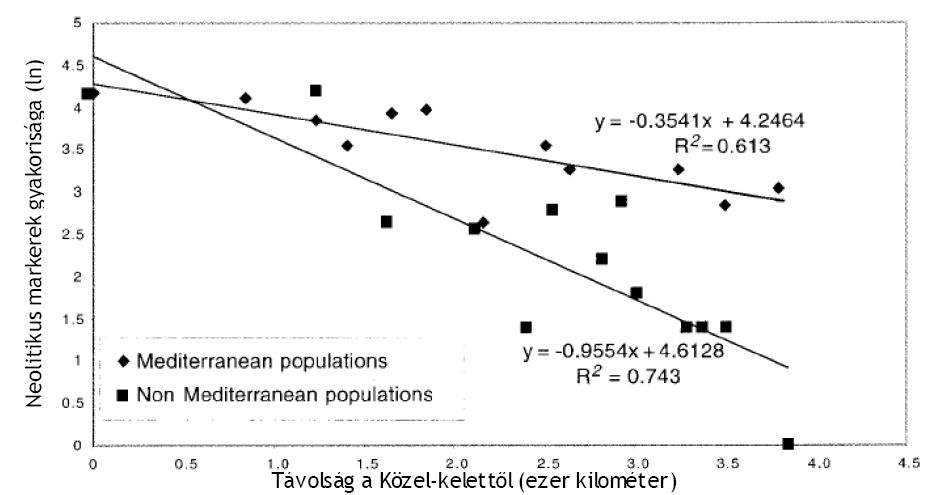
2. ábra Abszcissza: Az összes populáció távolsága ezer kilométerben a két közel-keleti populáció (libanoni és szíriai) átlagától. Ordináta: A neolitikus markerek (of Eu4, Eu9, Eu10 és Eu11) relatív gyakoriságának logaritmusa a mediterrán és nem mediterrán populációkban. A közel-keleti pont (X =0). (Az ábra forrása: Semino és mtársai 2000.)
A mediterrán populációk regressziós vonalának a lejtése jelentősen eltér a többi népességtől, jelezve, hogy a neolitikus földművelők szétszóródása elsősorban Dél-Európát érintette és kevésbé Közép-Európa térségét.
A megközelítőleg 4000 éves, Észak-Európa területén élő populációkra korlátozódó, újabb keletű uráli migrációt jelölő TAT és az M178 mutáció nem volt kimutatható Magyarországon, ahol mint köztudott az uráli nyelvcsalád egyik típusát beszélik.
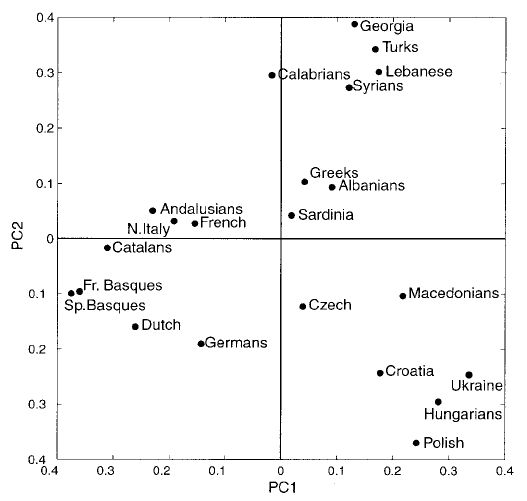
Az 1007 férfitól származó adatokból készített főkomponens analízis eredményét a 3. ábra mutatja be. Az ábrán megkülönböztethető három földrajzi-kulturális csoportosulás. Az első a baszkok és a nyugat európaiak, a második a közel-keletiek, a harmadik a kelet-európaiak Horvátországból, Ukrajnából, Magyarországból és Lengyelországból származó mintáit jeleníti meg. A földrajzi csoportoknak az elhelyezkedése lényegében megegyezik a fő jégkorszaki menedékhelyek illetve a földművelés expanziójának kiindulási régióival.
3. ábra A főkomponens analízis. Az első főkomponens (PC1) a variancia 46,24%-át, a második (PC2) a 34.69%-át teszi ki.
A európai anyai öröklődésű mitokondriális szekvencia vizsgálatok eredményei azt mutatják, hogy az anyai eredetű mitokondriumok 80%-a paleolotikumi eredetű, és csak 20%-a neolitikus örökség. Jelen vizsgálat adatai is megegyeznek ezzel a megfigyeléssel, mert az apai eredetű Eu4, E9, Eu10 és Eu11 összesen csak 22%-át adja az európai férfi nemi kromoszómáknak.
A korábbiakban kívülállóknak tartott szardíniaiak, baszkok és lappok alapjaiban azonos Y bináris komponenseket hordoznak, mint a többi európai népesség. A géngyakoriságaik alapján elfoglalt sajátos pozícióik a többi népességhez képest valószínűleg genetikai sodródás és izolációs folyamatok következményei lehetnek.
Az elemzés alapján szinte az összes európai férfi Y-kromoszóma mintázata belesorolható volt a tíz darab leszármazási vonal valamelyikébe. Ha röviden össze akarjuk foglalni a közlemény fő megállapításait, akkor az a következőképpen szólna: az európai génállomány jelentős része felső paleolitikumi eredetű (4. ábra), aminek földrajzi megoszlása azonban a jégkorszak leghidegebb periódusában, illetve azt követően megváltozott, átalakult, amikor Európa korábban a jég miatt lakatlanná vált régiói része újra benépesültek a kedvező irányú éghajlatváltozás következményeként. A későbbi neolitikus földművelők Y-kromoszómáinak szétszóródása elsősorban Dél-Európát, a Mediterráneum vidékét érintette és lényegesen kevésbé Európa északi térségét a vizsgálat szerint.
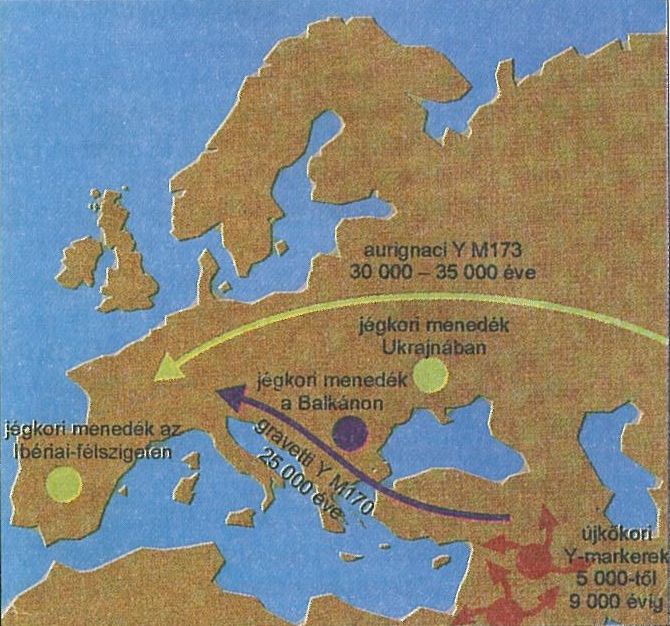
4. ábra A felső paleolitikumi fő vándorlási irányok és az általuk hozott Y-kromoszóma mutációk Semino és mtsi közleménye alapján. 40-35 ezer éve Ázsia felől érkeztek Európába az M173 mutáció (Eu18, zöld) vándorlási útvonala (aurignaci kultúra). A Közel-Keletről 20-25 ezer éve Dél és Közép-Európába bekerülő M170 mutációt (Eu7, bordó) útvonala (gravetti kultúra) (Az ábra forrása: Pásztor Erzsébet 2001.)
Források:
- SEMINO, O és mtársai 2001. The Genetic Legacy of Paleolithic Homo sapiens sapiens in Extant Europeans: A Y Chromosome Perspective. 10 NOVEMBER 2000 VOL 290 SCIENCE
- PÁSZTOR Erzsébet 2001. A férfiak európai útja. Élet és Tudomány 2001/2. 52-55. oldal
- BÉRES Judit 2003. A magyarországi népesség genetikai rokonsága. In. Hídvégi Egon (szerk.) Széphalom Könyvműhely. A genom 171-185. oldal
- GÁSPÁR Róbert 2007. A genetikai markerek és a magyar őstörténet. In Acta Historica Turiciensia XXIII. évfolyam 1. szám A Zürichi Magyar Történelmi Egyesület 54. sz. kiadványa, Huszadik Magyar Őstörténeti Találkozó Előadásai és Iratai Hódmezővásárhely 2007.
Ajánlott bejegyzések:
A bejegyzés trackback címe:
Kommentek:
A hozzászólások a vonatkozó jogszabályok értelmében felhasználói tartalomnak minősülnek, értük a szolgáltatás technikai üzemeltetője semmilyen felelősséget nem vállal, azokat nem ellenőrzi. Kifogás esetén forduljon a blog szerkesztőjéhez. Részletek a Felhasználási feltételekben és az adatvédelmi tájékoztatóban.